
Truth Journal
THE EFFECT OF SILENT THINKING ON THE CEREBRAL CORTEX
Sir John C. Eccles
1. Thinking
With the exception of the hard-core radical materialists there is general agreement on the existence of mental events such as thinking. Thinking is of course subjectively experienced and is not objectively identifiable in the way that we perceive the world around us through our senses. Popper has sharpened this distinction by his classification of the whole world of matter-energy as World I and the world of subjective experience as World 2. A test of such a division is provided by the analysis of such experiences as illusions and hallucinations, which, though interpreted by the experiencer as being in World 1, have to be recognized as being in World 2. For example, the apparently objective world of a dream experience has in fact no World I status, but is entirely in World 2. Of course, simultaneously with the dream, objective World I happenings can be recognized in the brain and even in the body of the dreamer, but they give no evidence of the content of the dream as recounted on awakening.There are wide ranges of mental events as illustrated in the World 2 list in Fig. I, but all can be subsumed under the general term "mind", which now is again in general philosophical use after its eclipse by Ryle's (1949) great cathartic book The Concept of Mind . The recovery is signalled by such books as The Existence of Mind (Beloff, 1962) and Content and Consciousness by Dennett (1969), where it is stated (p. 99) "The most central feature of mind, the 'phenomenon' that seems more than any other to be quintessentially 'mental' and non-physical is consciousness". In their book The Self and Its Brain (1977) Popper and Eccles give equal status to World I and World 2, a dualism as sharp as that of Descartes, and are specially concerned with the problem of interactionism .
Finally in his book The Nature of Mind Armstrong (1981), though still regarding himself as a physicalist, presents a most insightful account of consciousness and introspection stating (p. 68) that "Without introspective consciousness there would be little or no memory of the past history of the self--we would not be aware that we existed."
In Fig. I "thinking" is listed as one of the subjective experiences in World 2. It is a word covering an immense range of mental happenings. In order to limit philosophical discussion Beloff (1962, p. 98) defines thinking "as the activity of problem solving", but he immediately enlarges the scope by stating that a problem exists whenever some end is sought, but the means to that end has still to be discovered. That definition is too narrow for our present purpose where thinking is mental concentration that usually is on some particular learnt program or algorithm, but which also may be mental concentration on some anticipated sensory input. Thus we can limit our concept of thinking to mental concentration on some particular task. Philosophical discussion on the concept of thinking and the brain-mind problem will follow an account of the extraordinary findings on human brains engaged in thinking that is related to a variety of learned procedures. In this way it comes about that there is an internal generation of thinking in the absence of any signalling to the brain by sense organs.
2. Cerebral Events During Thinking
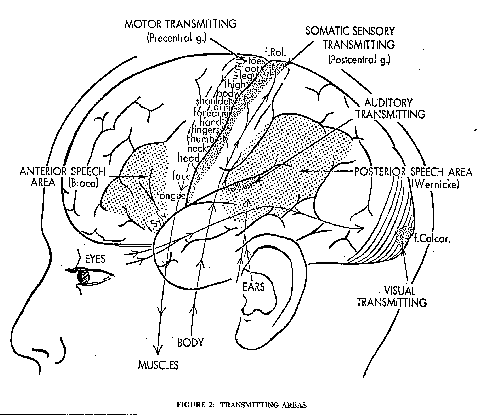
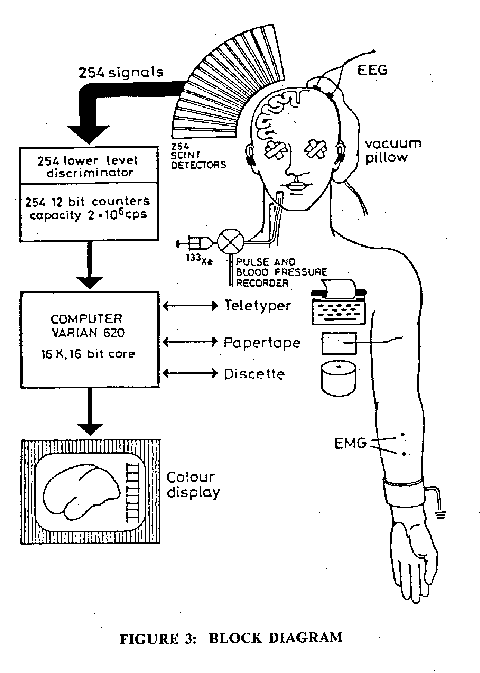
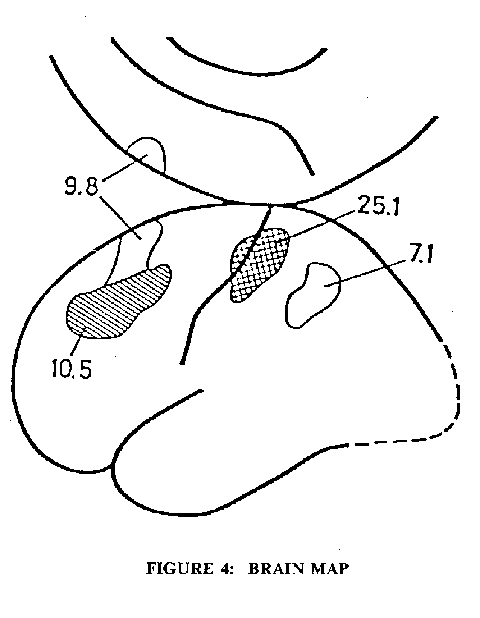
Fig. 2 shows the brain (Cerebral Cortex) in the head with the rather arbitrary division into the four lobes, frontal, parietal, temporal and occipital, with the primary visual, auditory, somato-sensory and motor areas indicated. This will serve as a basic cortical map for orientation of the areas activated in various types of silent thinking. Fig. 3 gives a diagrammatic representation of the extremely complicated procedures used for measuring the regional cerebral blood flow (rCBF) during thinking (Roland et al., 1980, Roland 1981; Roland and Friberg, 1985). With present techniques the spatial resolving power of the radioactive Xenon (133xe) technique is no better than I Ocm . The rCBF can be assumed to be a reliable indicator of the neuronal activity in that area. An assemblage of 254 collimator tubes over one cerebral hemisphere recorded that emission from the 133xe in the cerebral circulation for 45 seconds after the I second long injection of the solution into the internal carotid artery (Fig. 3). Fig. 3 indicates in a greatly simplified form the steps whereby there is an integration and synthesis of the data from the 254 assemblage into a rCBF map of the subjacent cerebral cortex. The technique necessarily involves the very coarse temporal grain of 45 to 60 seconds, which is the time for clearance of the 133xe from the cerebral blood flow. When determining the rCBF for some specific cerebral action, it is essential to allow for the background activity as indicated by the rCBF of the cerebral cortex when the subject is literally thinking of nothing. In the rCBF maps here illustrated this allowance has always been made, and moreover there has been synthesis of several experimental runs on the same or other subjects with calculation of the statistical significances of the percentage increases. It is remarkable that in all the diverse experimental tests no significant rCBF reduction of any area was ever observed.Fig. 4 illustrates a remarkable finding of Roland (1981) that, when the human subject was concentratedly attending to a finger on which just detectable touch stimuli were to be applied, there was an increase in the rCBF over the finger touch area of the postcentral gyrus of the cerebral cortex (Fig. 2) as well as in the mid-prefrontal area. These increases must have resulted from the mental attention because actually no touch was applied during the recording. Thus, Fig. 4 is a clear demonstration that the mental act of concentrated attention (thinking) can activate appropriate regions of the cerebral cortex. A similar finding occurs with attention to the lips in expectation of a touch, but of course the activated somatosensory area is now for the lips.
The effect of concentrated attention in causing an increased cerebral electrical response to finger touch has also been demonstrated by Desmedt and Robinson (1977). In a very ingenious investigation they discovered that, with touch to the attended finger, there was a large increase in the late N 140 and P 500 waves of the evoked potentials relative to controls with touch to unattended fingers. This may be correlated with the increased rCBF that attention produced in the finger area of Fig. 4. In both these investigations the concentrated attention or thinking was effecting selective neuronal responses.
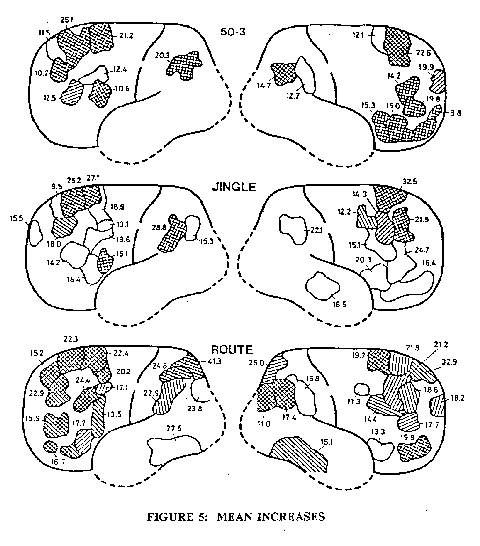
More recently Roland and Friberg (1985) have carried out a similar investigation on more complex thinking procedures. The subject had to learn these procedures by repeated rehearsal. Thus in all cases there is a background of instructional indoctrination, but it always ceased well before the experimental run with the injected 133xe. The subject is Iying with eyes and ears closed (cf. Fig. 3) and is at complete rest with no muscular movements except for quiet breathing. This is the background on which the silent thinking is superimposed. Three distinct types of silent thinking were investigated, the pooled results of each being displayed in Fig. 5. In order to avoid the development of automatic responses the initial training procedures were on a related but different paradigm from that presented to the subject some time (> 27 sec) before the actual experimental run.
In Fig. 5 the 50-3 frame shows the statistically significant increases when the subject was silently carrying out sequential subtractions of 3's from 50, the thinking procedure being 50-47-4441 -38, etc., continuing on below 2 to - I - -4, etc., until after the end of the recording of the rCBF at about 45 sec. During all that period the subject was undisturbed by the environment. The first disturbance after the rCBF recording period was by the investigator who asked the subject the last number reached by the subtraction procedure. It is remarkable that the significantly increased rCBF's were on both sides in the prefrontal lobe except for the angular gyrus in the parietal lobe with increases of 20.3 and 14.796. Correspondingly the clinical condition, acalculia, has been observed with bilateral destruction of the angular gyrus.
In Fig. 5 the "jingle" frame shows the rCBF increases when the silent thinking was concentrated on a task of jumping mentally to every second word of a well-known Danism nonsense word sequence or jingle which consists of a closer loop of 9 words. Again almost all of the activated cortical areas were in the frontal lobe. The area of the right temporal lobe with a 16.5% increase was uniquely involved in jingle thinking and had previously been implicated in discrimination of auditory inputs (Roland et al., 1981).
The lowest frame of Fig. 5 gives the rCBF's during a route-finding paradigm, which is designed to test the effects of silently thinking of sequential visual scenes. The subjects Iying silently with eyes sealed had to imagine what they saw when they walked out of their front door into the street and turned left, then right at the next corner, left at the next and so on. It involved an immense amount of silent visual thinking, and correspondingly the cortical rCBF map was more complex bilaterally and with the involvement of visual areas. Unfortunately the 133xe testing procedure gives little information from the occipital and inferior temporal lobes because the internal carotid artery is very little distributed to these areas.
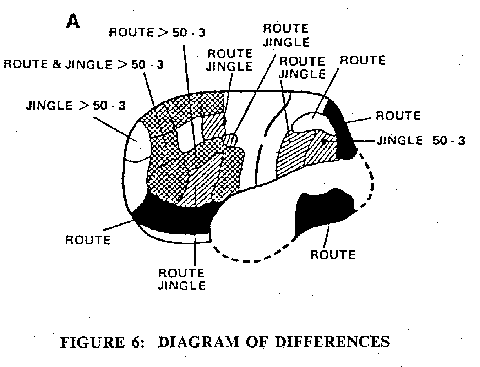
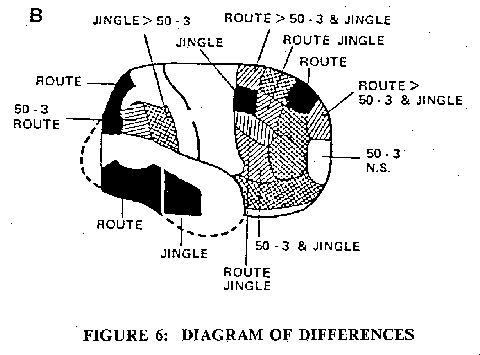
Despite the extreme differences in the types of silent thinking, number sequences, word sequences, visual sequences, Fig. 5 shows that in all three there was activation of the same cortical areas of both hemispheres, particularly in the superior and mid-frontal lobes. The similarities and differences are shown in the superimposed cortical maps of Fig. 6. Notable in Fig. 6 are the uniquely activated areas shown in black. For example the occipital superior and the posterior inferior temporal on both sides are the visual areas for route-finding, and the right mid-temporal is an auditory area for the jingle. The prefrontal lobe is most complexly involved and will require a finer grain of discrimination for elucidation in detail. There is an important negative finding in Fig. 6, namely that all these three types of silent thinking fail to generate significant increases in rCBF's in any of the primary or secondary motor and sensory cortical areas delineated in Fig. 2.
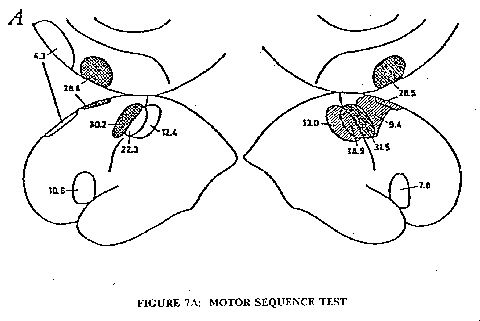
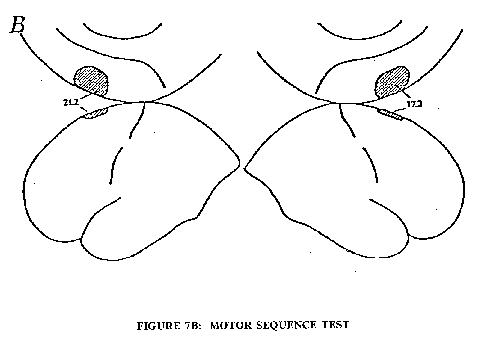
A quite different type of silent thinking was investigated by Roland et al. (1980) utilizing a sequential motor task, but with the same 133xe technique. In the initial investigation the subject continuously carried out a learned movement during the rCBF measurement. The particular example chosen was to touch rapidly the thumb by each finger in turn, two touches by finger 1, one by finger 2, 3 by finger 3 and 2 by finger 4, then reverse, 2 to 4, 3 to 3, I to 2 and 2 to I, and so on. The subject learns this motor sequence, as it is called, so that it is carried out without error, but it still requires concentrated thinking. In Fig. 7A the rCBF map shows large increases in the motor and sensory areas for fingers and thumb (cf. Fig. 2) on the contralateral side, which is in accord with the well known crossed innervation of limbs by the cerebral cortex. But also in Fig. 7A there is activation of an area that is largely on the medial side of the frontal cortex, and this activation of the supplementary motor area (SMA), as it is called, is bilateral and just about as strong as for the motor cortex. The primary role of the SMA in voluntary movement is disclosed in Fig. 7B. With prior instruction and training the subject is able to carry out the motor sequence test by silent thinking in the absence of any movement, which is checked by continuous electro-hyographic recording. Under such conditions the SMA is bilaterally activated on both sides by the internal programming, as it is called, and there is no significant increase in rCBF in any other cortical area. Fig. 7B thus indicates a very selective action by silent thinking of a learned motor movement.
Fig. 7 illustrates an important caveat that enters into all the rCBF maps. Since the silent thinking of the motor sequence activates only the SMA (Fig. 7B), it can be assumed that the additional activation with the motor sequence movements (Fig. 7A) is due to the excitatory action of the SMA neurones on the motor cortex. The further activation of the somatosensory area could be attributed to sensory feedback from the finger and thumb movement. Thus it is not necessary to assume that the thinking directly activated the areas in Fig. 7A additional to those in Fig. 7B. Similarly in Fig. 5 many of the activated areas could be secondary to a few that are primarily activated. In the three types of silent thinking it could be that the primary action is on areas of the superior prefrontal cortex on both sides (cf. Fig. 6) and these in turn cause the activation of the other areas, but there is as yet no experimental evidence for this as there is in Fig. 7. Possibly the primary action of the silent thinking in Fig. 4 is on the area of the mid-prefrontal cortex, which secondarily involves that area of the somatosensory cortex corresponding to the mental concentration.
3. The Mind-Brain Problem
The empirical findings illustrated in Figs. 4, 5, 6 and 7 have great philosophical significance in respect of the mind-brain problem. The mental processes of silent thinking evoke remarkable responses in the brain and these responses exhibit a topography that is different for different types of thinking. It is important to recognize that the 133xe technique can give only a crude spatio-temporal pattern and that much of the activity may be due to subsequent neuronal activation- but this must not obscure the significance of the discovery that the mental events of silent thinking occur in conjunction with specific patterns of response in the cerebral cortex. It was surprising to find that the silent thinking for the jingle and route-finding procedures evoked increases in the total blood flow to the brain which were in excess of that occurring in intense perceptual and motor tasks (Roland and Friberg, 1985, p. 1242). There is good justification for their conclusion that silent thinking is hard brain work! It has been an invariable finding that in the deepest mental relaxation the frontal lobes retain a level of rCBF higher than any other areas of the cortex. Possibly this is generated by all the random thinking in the awake state, which is sometimes referred to as day-dreaming (Ingvar, 1985).In general terms there are two theories about the relationship of mental events to neural events.
Firstly there is the explanation inherent in all monist-materialism including all the varieties of parallelism, panpsychism, epiphenomenalism and the now fashionable identity theory. The existence of mental events is not denied, but they are given a subsidiary role in the performance and experience of a human person, which is entirely brain controlled. For example in the identity theory the mental events are somehow regarded as 'identical' with neural events of a special kind in the highest levels of the brain as succinctly expressed by Feigl (1967, pp. 79 and 90): "The identity thesis which I wish to clarify and to defend asserts that the states of direct experience which conscious human beings 'live through', and those which we confidently ascribe to some of the higher animals, are identical with certain (presumably configurational) aspects of the neural processes in those organisms . . . processes in the central nervous system, perhaps especially in the cerebral cortex.... The neuro-physiological concepts refer to complicated highly ramified patterns of neuron discharges".
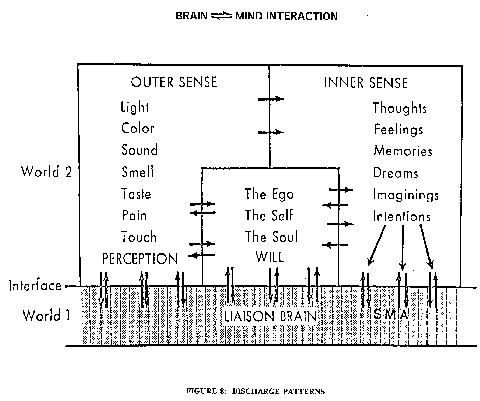
As Popper (Popper and Eccles, 1977, p. 51) points out, all materialist theories of the mind: "assert that the physical world (World I ) is self-contained or closed . . . This physicalist principle of the closedness of the physical World I is of decisive importance . . . as the characteristic principle of physicalism or materialism". Secondly, there is the dualist-interactionism explanation which has been specially developed for the self-conscious mind and human brains. It is proposed that, superimposed upon the neural machinery in all its performance, there are at certain sites of the cerebral hemispheres (the so-called liaison areas) effective interactions with the self-conscious mind, both in receiving and in giving (Fig. 8).
4. A New Hypothesis of Mind-Brain Interaction
The materialist critics argue that insuperable difficulties are encountered by the hypothesis that immaterial mental events such as thinking can act in any way on material structures such as neurons of the cerebral cortex, as is dia8rammed in Fig. 8. Such a presumed action is alleged to be incompatible with the conservation laws of physics, in particular of the First Law of Thermodynamics. This objection would certainly be sustained by 19th century physicists and by neuroscientists and philosophers who are still ideologically in the physics of the 19th century, not recognizing the revolution wrought by quantum physicists in the 20th century. Unfortunately it is rare for a quantum physicist to dare an intrusion into the brain-mind problem. But in a recent book the quantum physicist Margenau (1984) makes a fundamental contribution. It is a remarkable transformation from 19th century physics to be told (p. 22): "that some fields, such as the probability field of quantum mechanics carry neither energy nor matter." He goes on (p. 96) to state: "In very complicated physical systems such as the brain, the neurons and sense organs, whose constituents are small enough to be governed by probabilistic quantum laws, the physical organ is always poised for a multitude of possible changes, each with a definite probability; if one change takes place that requires energy, or more or less energy than another, the intricate organism furnishes it automatically. Hence, even if the mind has anything to do with the change, that is if there is a mind-body interaction, the mind would not be called upon to furnish energy." In summary (p. 97) Margenau states that: "The mind may be regarded as a field in the accepted physical sense of the term. But it is a nonmaterial field; its closest analogue is perhaps a probability field. It cannot be compared with the simpler nonmaterial fields that require the presence of matter (e.g. gravity) nor does it necessarily have a definite position in space. And so far as present evidence goes it is not an energy field in any physical sense, nor is it required to contain energy in order to account for all known phenomena in which mind interacts with brain".In formulating more precisely the dualist hypothesis of mind-brain interaction, the initial statement is that the whole world of mental events (World 2) has an existence as autonomous as the world of matter-energy (World I ) (Fig. 1). The present interactionist hypothesis does not relate to these ontological problems, but merely to the mode of action of mental events on neural events, that is of the nature of the downward arrows across the frontier in Fig. 8. Following Margenau (1984) the hypothesis is that mind-brain interaction is analogous to a probability field of quantum mechanics, which has neither mass nor energy, yet can cause effective action at microsites. More specifically it is proposed that the mental concentration involved in intentions (Figs. 7, 8) or attention (Fig. 4) or planned thinking (Figs. 5, 6) can cause neural events by a process analogous to the probability fields of quantum mechanics.
We can ask: what neural events could be appropriate recipients for mental fields that are analogous to quantal probability fields? Already we may have the answer in recent discoveries on the nature of the synaptic mechanism whereby one nerve cell communicates with another.
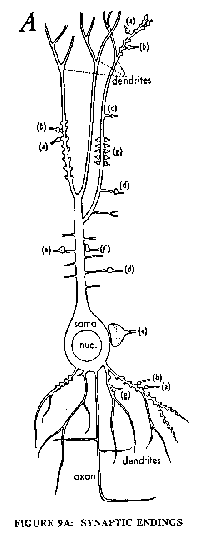
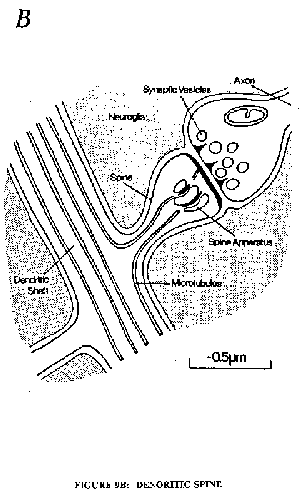
Fig. 9A is a diagrammatic representation of a nerve cell of the cerebral cortex showing the soma with a long apical dendrite studded with spines on each of which there is a synapse from a nerve terminal derived from some other nerve cell. There are about 10,000 spine synapses on each pyramidal cell. Fig. 9B gives a diagram of such a synapse showing the nerve fibre expanded to a terminal bouton that makes a close contact with a special membrane thickening of the spine. In the bouton are numerous vesicles each of which contains 5,000 to 10,000 molecules of the specific synaptic transmitter substance, which is glutamate or aspartate for the great majority of excitatory boutons in the cerebral cortex. Some synaptic vesicles are in close contact with the presynaptic membrane confronting the postsynaptic membrane across the extremely narrow synaptic cleft. These synaptic vesicles appear to be arr Inged between dense projections.
Further structural analysis particularly by the freeze-fracture technique of Akert and associates (1972, 1975) has led to the construction of a diagram of an idealized spine synapse (Fig. 10), which is shown in perspective with partial excisions to reveal the deeper structures. The relatively loose arrangement of synaptic vesicles and presynaptic dense projections (Fig. 9B) is shown in Fig. 10 as the precise packing illustrated in the inset on the left, with the synaptic vesicles in hexagonal array packaged between the presynaptic dense projections in triangular array. This composite structure is termed a presynaptic vesicular grid and it can be regarded as having paracrystalline properties (Akert et al., 1975; Triller and Korn, 1982). The boutons of brain synapses have a single presynaptic vesicular grid, as in indicated in Figs. 9B and 10.
There are only approximate figures for the number of synaptic vesicles incorporated in a presynaptic vesicular grid. The usual number appears to be 30 to 50 from the illustrations of Akert et al. (1972, 1975). Triller and Korn (1982) give the number as 44-83 for the boutons on Mauthner cells. Thus only a very small proportion of the synaptic vesicles of a bouton (about 20,000) are embedded in the firing zone of the presynaptic vesicular grid. The remainder are loosely arranged in the interior of the bouton, as is partly shown in Figs 9B and 10.
Fig. l l A well illustrates the packaging of transmitter molecules into a synaptic vesicle, its movement up to the presynaptic vesicular grid with locking into a presynaptic density of the presynaptic grid. Finally in Fig.l IB there is apposition to the presynaptic membrane under the influence of Ca2+ ions and the total release of the transmitter molecules into the synaptic cleft. The very close contact of the vesicle to the presynaptic membrane (Fig.l IB2) is also depicted in the left of Fig.10. with the two little bulges and one vesicle apparently ready to discharge, while to the right of Fig.10, after the vesicles and the dense projections have been stripped off, the vesicle attachment sites, V.A.S., are seen in hexagonal array, as also in the inset diagram to the right.
By precise analysis of the postsynaptic potentials generated when a presynaptic impulse activates a single bouton (Fig. 9A, 10), it has been shown that the vesicular emission from a bouton (exocytosis) is probabilistic and below unity - the usual probability being 0.5 or less (Jack et al., 1981 A; Korn and Faber, 1986). This probability can be varied up or down according to circumstances. The presynaptic vesicular grid must have some subtle functional organization in controlling exocytosis of the embedded vesicles. It is proposed that mental events such as intention (Fig. 7), attention (Fig. 4) or silent thinking (Figs. 5, 6), acting in a manner analogous to a probability field of quantum mechanics, change the probability of synaptic vesicular emission in the manner suggested by Margenau (1984, pp. 96, 97). It is not postulated that the firing of neurons is the target of the mental events, but merely that this firing is modified by alterations of the probabilities of quantal emission of those synapses that are engaged in actively exciting them. This is an important limitation in the target for events such as the varieties of mental concentration in Figs. 4, 5, 6, 7.
The first question that can be raised concerns the magnitude of the effect that could be produced by a probability wave of quantum mechanics. Is the mass of the synaptic vesicle so great that it lies outside the range of the uncertainty principle of Heisenberg? Margenau (p. 384) adapts the usual uncertainty equation for this calculation of non-atomic situations.
However, in Fig. I IB it can be seen that the mass critically involved in the exocytosis is much less than the mass of a vesicle which is already in position (cf . Figs. 10, 11 B2). All that is required is the displacement of a small area of the double membrane. This area as depicted in Figs. IIB2 to llB3 would be no more than 10 nm thick, and if it was 10 nm by 10 nm in area it would have a mass of only 1018 g. If the uncertainty of the position, x, of the contact site is taken to be 1 nm, then v, the uncertainty of the velocity, comes out at 10 nm in 0, I msec, which is of the order expected for the exocytotic opening seen in Fig. 11 B3. Since the vesicle is already in position (Fig. I lB2) in the presynaptic vesicular grid (Fig. 10), the exocytosis is not dependent on movement through a viscous medium. The postulated mental influence would do no more than alter the probability of emission of a vesicle already in apposition.
It can be concluded that calculations on the basis of the Heisenberg uncertainty principle show that the probabilistic emission of a vesicle from the presynaptic vesicular grid could conceivably be modified by a mental intention acting analogously to a quantal probability field.
The second question raises the order of magnitude of the effect, which is merely a change in probability of emission of a single vesicle (Figs. 10, 11). This is many orders of magnitude too small for modifying the patterns of neuronal activity even in small areas of the brain. However, there are many thousands of similar boutons on a pyramidal cell of the cerebral cortex (Fig. 9A). The hypothesis is that the probability field of the mental intention is widely distributed not only to the synapses on that neurone, but also to the synapses of a multitude of other neurones with similar functions.
According to the microsite hypothesis the presynaptic vesicular grid provided the change for the mental intention to change by choice the probability of its synaptic emission. This would be happening over the whole ensemble of spine synapses that are activated at that time, probably even thousands, since there are about 10,000 on a single cortical pyramidal cell (Szentagothai, 1978A). It would be expected that a mental influence analogous to a probability field would exert a global influence on the synapses of an appropriate neurone modifying up or down the probabilities of vesicular emission by incoming impulses. So the reliability of mental intention is derived from integration of the chance happenings at the multitude of presynaptic vesicular grids on that neurone.
As already mentioned, Margenau (1984) makes the prescient statement that, in the postulated action of a mental event on a neuronal event at microsites, there need be no energy requirement. "If one change takes place that requires energy . . . the intricate organism furnishes it automatically". Our attention should be focussed on the exocytosis of a vesicle as indicated in Fig. 11 B2 to 3. Though the displacement of the apposition membrane may require a minute energy, the exocytotic release of the stored transmitter molecules in Fig. 11 B3 could be a source of energy. Thus on the micro-site hypothesis the probability of exocytosis in response to a nerve impulse may be altered by a mental event without an energy requirement. The conservation laws would remain inviolate.
5. Testing of Mind-Brain Theories
We can raise the question whether there could be experimental testing of predictions from the dualist-interactionism hypothesis (Fig. 8) on the one hand and the identity hypothesis on the other. A simple diagram (Fig. 12A) embodies the essential features of the identity hypothesis. In accord with Feigl (1967) mental-neural identity occurs only for neurons or neuron systems at a high level of the brain, especially in the cerebral cortex. These neurons can be called mental-neural event (MNE) neurons, whereas other neurons in the brain, and in particular neurons on the input and output pathways, would be no more than simple neural event (NE) neurons, as in the diagram (Fig. 12A). It would be predicted from the identity hypothesis that MNE neurons would be distinctive because in special circumstances their firing would be in unison (the identity) with mental events. But of course this firing would be in response to inputs from other neurons, MNE or NE, and is in no way determined by or modified by the mental events. This is the closedness of the physical world referred to above (Popper and Eccles. 1977).However, the diagram of Fig. 12A is challenged by the evidence that internally Benerated thoughts strongly excite neurones in special areas of the cerebral cortex (Figs. 4, 5, 6, 7, 8). These findings require the diagrammatic addition of an input from mental events (ME) per se as is shown by the additional arrows in the diagram (Fig. 12B). The firing of MNE neurons would exhibit a response that is different from what it would be in the absence of the mental events of attention, silent thinking or intention as is shown in Figs. 4, 5, 6, 7. Fig. 12B is a diagrammatic representation of dualist interactionism.
We can return to Fig. I with the juxtaposed World I and World 2 each with a fundamental primacy, something that we have to accept as given, as two distinct orders of existence. In that context it can be appreciated that this lecture describes an attempt to show how microsites in the brain could have transcendental properties of being channels of communication between these two completely disparate existences. The philosophical implications could be far reaching if it comes to be accepted that mental events can effectively act on the brain as indicated in Fips. 4, 5, 6, 7, 8. We all think and act as if we have at least some control and responsibility for our actions, especially our linguistic expressions, but reductionist critics have insisted that this must be an illusion since it is contrary to the conservation laws of physics. We are now free to reject these criticisms.
It may seem beyond belief that the immense neuronal activity demonstrated in Fig. 5 can be generated by internal thinking that initially merely modified the probability of quantal emission from synaptic boutons. However, an essential component of the microsite hypothesis is that the mental event of thinking is widely distributed to the boutons of one pyramidal cell (cf. Fig. 9A) taking advantage of the design feature of an immense number of boutons (^ 10,000) on one such cell. Hence there is opportunity for great amplification at the primary site of action with the consequent bursts of impulse discharges such as was observed with SMA neurones in responses to voluntary movements of monkeys (Brinkman and Porter, 1979).
From the postulated net zero energy influence on the probability of vesicular emission at one microsite, there are three factors giving an immense amplification: the many microsites on one pyramidal cell; the many pyramidal cells subjected to the same mental influence - a functional cluster; the spreading excitation from the excited pyramidal cells by their axon collaterals and the excitatory stellate cells in the adjacent cortex. Hence we can account for the high level of "brainwork" observed by Roland and Friberg (1985).
On the microsite hypothesis it would be predicted that the interaction with mental events would be reduced to zero when the presynaptic firing background was reduced to zero. Loss of consciousness would occur, and be irreversible unless there would be revival to a considerable degree of the impulse discharges in the cerebral cortex. An example is "vigil comaz that supervenes when brain injury to the mid-brain turns off the reticular activating system (Hassler, 1978; Eccles, 1980, p. 160). In fact the principal role of the reticular activating system may be to provide a background of excitatory synaptic actions on neurons of the cerebral cortex with an immense array of probabilistic vesicular emissions that are targets for the quantal probabilistic fields of mental influences.
6. Summary Statements on the Brain-Mind Problem
A general observation is that hitherto all hypotheses attempt to give some explanation of how conscious experiences derive from or relate to neural events in the active cerebral cortex, as was done by Feigl (1967) in Section 5. Sperry (1976) proposed that mental events are holistic configurational properties of the brain process. Mountcastle (1978) developed the concept of distributed systems which are "composed of large numbers of modular elements linked together in echeloned parallel and serial arrangments", and are thought to provide an objective mechanism of conscious awareness. Edelman (1978) suggested that: "the brain processes sensory signals and its own stored information upon this selective base in a phasic (cyclic) and re-entrant manner that is capable of generating the necessary conditions for conscious states". Szentagothai (1978B) suggested that "dynamic patterns" offer "superstructures" and might be helpful to give a scientific explanation on the higher functions of the brain including even consciousness. Eccles (1982) suggested that "the mental influence is exerted on an extremely complex dynamic system of interacting neurons."The extreme alternative to these "nebular" hypotheses is now proposed, namely that the essential locus of the action of nonmaterial mental events on the brain is at individual microsites, the presynaptic vesicular grids of the boutons, each of which operates in a probabilistic manner in the release of a single vesicle in response to a presynaptic impulse. It is this probability that is assumed to be modified by a mental influence acting analogously to a quantal probability field in the manner described above. The mechanism by which effective action at microsites becomes amplified by conventional neuro-circuitry will be dependent on the complex circuits envisioned, for example, by Feigl (1967), Sperry (1976). Mountcastle (1978), Edelman (1978), Szentagothai (1978B) and Eccles (1982). The microsite hypothesis can be proposed as a tentative beginning of a scientific study of the reflective loop proposed by Creutzfelot (1979) as opening up the independent symbolic world of the mind, which is the World 2 of Popper and Eccles (1977). In contrast to the "nebular" hypotheses it offers a unique challenge to molecular neurobiology.
LEGENDS
Fig. 1: Tabular representation of the contents of the three worlds that comprise everything in existence and in experience.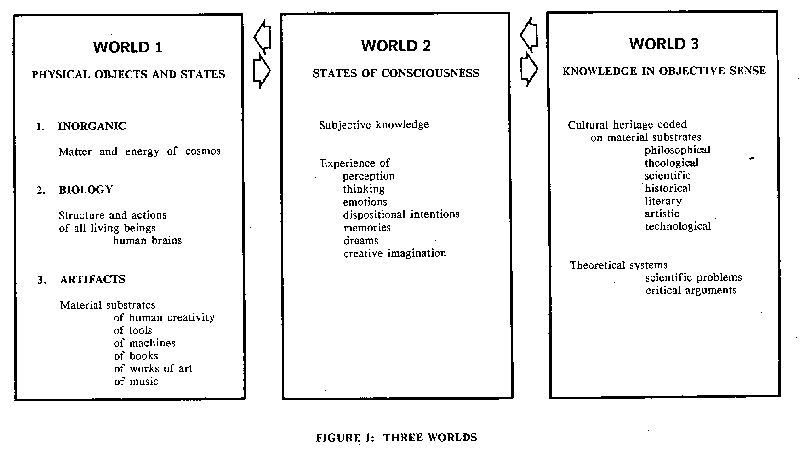{kind=link}
Fig. 2: The motor and sensory transmitting areas of the cerebral cortex. The approximate map of the motor transmitting areas is shown in the precentral gyrus, while the somatic sensory receiving areas are in a similar map in the postcentral gyrus. Actually the toes, foot and leg should be represented over the top of the medial surface. Other primary sensory areas shown are the visual and auditory, but they are largely in areas screened from this lateral view. The frontal, parietal, occipital and temporal lobes are indicated. Also shown are the speech areas of Broca and Wernicke.
{kind=link}
Fig. 3: Block diagram of the equipment and the principles of the method. The head is fixed to the collimator by a vacuum pillow. The 254 collimator tubes are arranged radially in a 50 mm-thick spherical lead segment. The spatial resolving power of the camera when used for rCBF measurements is one channel. The center-tocenter distance of 2 adjacent collimator tubes is 10 mm. Signals from the 254 cortical regions are processed on-line, the isotope clearance curves, the rCBF values, and background radioactivity can be recorded in the 3 different formats for further processing or displayed on the TV screen (Roland et al., 1980).
{kind=link}
Fig. 4: Mean increase of rCBF in percent during pure selective somatosensory attention; that is, somatosensory latent sensing without peripheral stimulation. The size and location of each focus shown is the geometrical average of the individual focus. Each individual focus has been transferred to a brain map of standard dimensions with a proportional stereotaxic system. The crosshatched areas have an increase of rCBF significant at the 0,0005 level (Student's test, one-sided significance level). For the other areas shown the rCBF increase is significant at the 0,05 level. Eight subjects. (Roland, 1981).
{kind=link}
Fig. 5: Mean increases of rCBF in percent and their average distribution in cerebral cortex under three different conditions of silent thinking as described in the text. Left hemisphere six subjects, right hemisphere five subjects. Cross-hatched areas have rCBF increases significant at the 0,005 level. With hatched areas P < 0,01 and with outlined areas P < 0,05 (Roland and Friberg, 1985).
{kind=link}
Fig. 6: Diagram of differences in cortical activations during three different types of thinking. Areas that were specifically and statistically significantly activated in one type of thinking only are shown black. Areas activated by two types of thinking only, are shown hatched.
A: Left hemisphere;
{kind=link}
B: Right hemisphere (Roland and Friberg, 1985).Fig. 7:
{kind=link}
A. Mean increase of the rCBF in percent during the motor-sequence test performed with the contralateral hand, corrected for diffuse increase of the blood flow. Cross-hatched areas have an increase of rCBF significant at the 0,0005 level. Hatched areas have an increase of rCBF significant at the 0,005 level, for other areas shown the rCBF increase is significant at the level 0,05. Left: left hemisphere, five subjects. Right: right hemisphere, 10 subjects.
{kind=link}
B. Mean increase of rCBF in percent during internal programming of the motor-sequence test, values corrected for diffuse increase of the blood flow. Left: left hemisphere, three subjects; Right: right hemisphere, five subjects (Roland et al., 1980).Fig. 8: Illustration of the discharge patterns of a neurone associated with flexion of the elbow during the lever pull for both the right (A) and left hand (C). B is the periresponse time histogram demonstrating the EMG activity of a representative elbow flexor, m. brachioradialia, in the right arm during the same 20 pulls as those in A, and shows that the neurone increased its discharge well before EMG activity increased. This was the case for the majority of neurones in which the discharge pattern could be compared with EMG changes (Brinkman and Porter, 1979).
{kind=link}
{kind=link}
Fig. 9:
A. Synaptic endings on neurons. A drawing of a hippocampal pyramidal cell to illustrate the diversity of synaptic endings on the different zones of the apical and basal dendrites, and the inhibitory synaptic endings on the soma. The various types of synapses marked by the letters A to G are shown in higher magnification to the right (From Hamlyn, 1962).
{kind=link}
B. Drawing of an synapse on a denoritic spine. The bouton contains synaptic vesicles and dense projections on the presynaptic membrane (Gray, 1982).Fig. 10: Schema of the mammalian central synapse. The active zone is formed by presynaptic dense projections (az). The postsynaptic aggregation of intramembraneous particles is restricted to the area facing the active zone. sv = synaptic vesicles, pa = particle aggregations on postsynaptic membrane (postsyn.). Note synaptic vesicles (sv) in hexagonal array, as is well seen in the upper left inset, and the vesicle attachment sites (vas) in the right inset. Further description in text. (Akert et al., 1975).
{kind=link}
{kind=link}
Fig. 11: Stages of synaptic vesicle development, movement and exocytosis.
A. The three steps involved in filling a vesicle with transmitter and bringing it to attachment to a presynaptic dense projection of triangular shape.
{kind=link}
B. Stages of exocytosis with release of transmitter into the synaptic cleft depicting the essential role of Ca2+ input from the synaptic cleft (Kelly et al., 1979).Fig. 12: Diagrams of mind-brain theories.
{kind=link}
A. The identity theory.
{kind=link}
B. Dualist-interactionism. Assemblages of neurons are shown by circles. NE represents the conventional neurons which respond only to neural events. MNE are neurons that are associated with both mental and neural events and are grouped in a larger circle representing the higher nervous system. In B, ME arrows represent mental influences acting on the neural population that is associated both with mental and neural events. All other arrows in A and B represent the ordinary lines of neural communication which are shown in reciprocal action.
{kind=link}
REFERENCES
Akert, K., Peper, K. & C. Sandri 1975 Structural organization of motor end plate and central synapses. In: Cholinergic Mechanisms (ed. P. G. Waser), pp. 43-57. New York: Raven Press.Akert, K. Pfenninger, K., Sandri, C. & H. Moor 1972 Freeze etching and cytochemistry of vesicles and membrane complexes in synapses of the central nervous system. In: Structure and Function of Synapses (ed. G. P. Papas & D. F. Purpura), pp. 67-86. New York: Raven Press.
Armstrong, D. M. 1981 The Nature of Mind. Cornell University Press, Ithaca, N. Y.
Beloff, J. 1962 The Existence of Mind. MacGibson and Kee, London.
Brinkman, C. & R. Porter 1979 Supplementary motor area in the monkey: activity of neurons during performance of a learned motor task. J. Neurophysiol. 42, 861-709.
Creutzfeldt, O. D. 1979 Neurophysiological mechanisms and consciousness. In: Brain and Mind . Ciba Foundation Series 69, pp. 217-233. Amsterdam: Elsevier-North Holland. Dennett, D. C. 1969 Content and Consciousness. Humanities Press, New York.
Desmedt, J. E. & D. Robertson 1977 Differential enhancement of early and late components of the cerebral somatosensory evoked potentials during forced-paced cognitive tasks in man. J. Physiol. 271, 761-782.
Eccles, J. C. 1980 The Human Psyche. Springer Internat.: Berlin, Heidelberg, New York.
Eccles, J. C. 1982 How the self acts on the brain. Psychoneuroendocrinology 7, 271-283.
Edelman, G. M. 1978 Group selection and phasic reentrant signalling: A theory of higher brain function. In: The Mindful Brain, pp. 51-100. MIT Press: Cambridge, Mass.
Feigl, H. 1967 The "Mental" and the "Physical". Minneapolis, Minn.: University of Minnesota Press.
Gray, E. G. 1982 Rehabilitating the denoritic spine. Trends in Neurosciences 5, 56.
Hamlyn, L. H. 1962 An electron microscope study of pyramidal neurons in the Ammon's Horn of the rabbit. J. Anat. 97, 189-201.
Hassler, R. 1978 Interaction of reticular activating system for vigilance and the truncothalamic and pallidal systems for directing awareness and attention under striatal control. In: Cerebral Correlates of Conscious Experience. (ed. P. A. Buser & A. RougeulBuser) Amsterdam: Elsevier-North Holland, pp. 110-129.
Ingvar, D. H. 1985 "Memory of the future". An essay on the temporal organization of conscious awareness. Human Neurobiology 4, pp. 127-136.
Jack, J. J. B ., Redman, S. J. & K. Wong 1981 The components of synaptic potentials evoked in cat spinal motoneurones by impulses in single group IA afferents. J. Physiol. 321, 65-96.
Kelly, R. B., Deutsch, J. W., Carlson, S. S. & J. A. Wagner 1979 Biochemistry of neurotransmitter release. Ann. Rev. Neurosci. 2, 399-446.
Korn, H. & D. S. Faber 1986 Regulation and sugnificance of probabilistic release mechanisms at central synapses. In: New Insights into Synaptic Function (ed. G. M. Edelman, W. E. Gall & W. M. Cowan). New York: Neurosciences Research Foundation Inc., J. Wiley & Sons Inc.
Margenau, H. 1977 The Nature of Physical Reality. Woodbridge, Conn.: Ox Bow Press.
Margenau, H. 1984 The Miracle of Existence. Wooc bridge, Conn.: Ox Bow Press.
Montcastle, V. B. 1978 An organizing principle for cerebral function: the unit module and the distributed system. In: The Mindful Brain pp. 7-50. Cambridge (Mass.): MIT Press.
Popper, K. R. & J. C. Eccles 1977 The Self and Its Brain. Springer Internat.: Berlin, Heidelberg, New York.
Roland, P. E. 1981 Somatotopical tuning of postcentral gyrus during focal attention in man. A regional cerebral blood flow study. J. Neurophysiol. 46, 744-754.
Roland, P. E. & Friberg, L. 1985 Localization in cortical areas activated by thinking. J.
Neurophysiol. 53, 1219-1243.
Roland, P. E., Larsen, B., Lassen, N. A. & Skinhj 1980 Supplementary motor area and other cortical areas in organization of voluntary movements in man. J. Neurophysiol. 43, 118-136.
Roland, P. E., Skinhj, E. & Lassen, N. A. 1981 Focal activation of human cerebral cortex during auditory discrimination. J. Neurophysiol. 45, 1139-1151.
Ryle, G. The Concept of Mind. 1949. London: Hutchinson.
Sperry, R. W. 1976 Mental phenomena as causal determinants in brain function. In: Consciousness of the Brain. (ed. G. G. Globus, G. Maxwell & I. Savondik) pp. 163-177 New York: Plenum.
Szentagothai, J. 1978A The neuron network of the cerebral cortex. A functional interpretation. Proc. Roy. Soc. Lond. B 201, 219-248.
Szentagothai, J. 1978B The local neuronal apparatus of the cerebral cortex. In: Cerebral Correlates of Conscious Experience. (ed. P. Buser & A. Rongeul Buser) pp. 131-138, Amsterdam: Elsevier.
Triller, A. & H. Korn, 1982 Transmission at a central inhibitory synapse. 111. Ultrastructure of physiologically identified and stained terminals. J. Neurophysiol. 48, 708-736.
 Printer-Friendly Version See
Related Resources
Printer-Friendly Version See
Related Resources
copyright (c) 1995-1999 Leadership U. All rights reserved.
This site is part of the Telling the Truth Project.
Updated: 11 June 1997